经络穴位的敏化特性
疾病时,病变器官或组织引起体表特定部位的腧穴出现牵涉性敏化现象,是近些年穴位本态研究的重要进展。朱兵课题组在全国20余家医院对20种内脏疾病的患者进行了体表敏感点探查,观察到内脏疾病引起体表敏感点或区与穴位分布存在较高的重叠性,与非敏化点相比,疾病引起的体表敏化点机械痛阈值显著下降。有研究者对内脏疾病引起的体表敏化点分布与穴位定位进行了统计,结果显示,热敏点与穴位的重合率为48.76%,激痛点及压痛点重合率为71%~92% 。感觉异常是探寻敏化穴位的主要依据,敏化穴位局部常出现痛、热、痒等感觉敏化改变,或伴有局部的皮肤形态改变,如皮疹、结节等表现,其中尤以机械痛觉敏化最为常见,这也完全契合《灵枢·经筋》中“以痛为输”的取穴理念。
同时,穴位敏化时会出现范围大小及功能的改变,即功能的可塑性。
如Rong等在大鼠结肠注射芥子油后,观察到“足三里”“上巨虚”穴出现范围扩大,同时针刺、艾灸“足三里”“上巨虚”穴对脊髓背角广动力神经元的激活效应显著增加;许建峰等观察到急性肠黏膜损伤能够引起大鼠“膝前”“足三里-上巨虚”穴区出现敏化,刺激上述敏化穴诱发坐骨神经干C类纤维动作电位发放的阈值较非敏化部位显著降低,同时诱发的C类纤维放电数目较非敏化区显著增加。李武等在大鼠“天枢”穴注射芥子油模拟穴位敏化,以空肠运动为评价指标,观察到“天枢”模拟敏化后,能够直接抑制空肠运动,同时电针刺激芥子油诱导的敏化“天枢”穴对空肠运动的抑制效应较敏化前显著增强,提示电针刺激敏化穴对内脏功能能够产生更佳的调节效应。
由于体表、内脏的信息以及不同形式刺激产生的信息编码均由感受器响应并传递,因此,从负责响应或传递感觉信息的感受器层面,开展穴位敏化及针灸效应的神经生物学机制研究,是揭示穴位敏化及其功能可塑性的重要切入点。
伤害感受器分布及功能
伤害感受器是指分布于皮肤、肌肉、关节、内脏中的游离感觉终末,其胞体位于DRG或三叉神经节,能够选择性地对伤害性刺激或潜在损伤组织的刺激产生反应,并负责将信息传递至脊髓背角或脊髓上水平。敏化及功能可塑性,是伤害感受器的最主要生理特性,主要有两种形式:一种为出现在感受器游离感觉终末的自发性敏化,主要表现为激活阈值降低,兴奋性的膜电位水平增加;另一种为脊髓背角神经元的wind-up现象,主要表现为神经元的放电频率增加、时间延长。这两种形式分别在外周及中枢水平介导了感觉的敏化。
如前所述,穴位敏化时能够观察到穴位出现机械痛、热痛、压痛等痛觉敏化现象,以及范围及功能的可塑性变化,这些变化均与局部的伤害感受器激活相关。皮肤中的伤害感受器主要分为两种亚型,均在穴区有大量的分布,一种是来自中直径神经元、有髓鞘的Aδ类伤害感受器,主要对高强度的机械和温度刺激响应,与快痛产生相关;另一种是来自小直径神经元、无髓鞘的C类伤害感受器,即CPN,与慢痛、继发痛的产生相关,可被各种伤害性刺激如机械、化学、热刺激激活。根据是否能够分泌神经肽,C类感受器又可以划分为肽能(分布广泛,在各种组织以及皮肤深层分布)与非肽能(与皮肤相关,主要分布在表皮)两种类型。目前研究认为,皮肤中分布最多的伤害感受器是CPN。
穴位敏化主要与C类伤害感受器激活相关
近些年的研究表明,C类伤害感受器的激活与穴位敏化的发生最为相关。如前述提到的动物内脏病变时进行伊文氏蓝实验,能够在体表观察到与穴位分布具有重叠性的蓝色的血浆蛋白渗出点,而这些渗出点的出现与局部的C类伤害感受器的激活密切相关。通过动物实验,目前已经明确C类伤害感受器参与穴位敏化。
如He等以胃黏膜损伤模型大鼠为研究对象,通过免疫荧光实验观察到敏化穴位皮肤中标记C类感受器的降钙素基因相关肽(CGRP)、P物质(SP)较非敏化穴显著增加;Zhang等以膝关节炎模型大鼠为研究对象,观察到膝关节炎引起“犊鼻”穴敏化;通过逆向示踪技术显示支配膝关节“犊鼻”穴的DRG中C类和Aδ神经元,并采用离体膜片钳方法记录C类和Aδ神经元的动作电位,观察到”犊鼻”穴敏化后,支配“犊鼻”穴的C类伤害性神经元产生动作电位的电流阈值显著降低,产生的超极化电流(Ih)水平及传导速度显著增加,而对Aδ类神经元的活动无显著影响;同时,他们进一步观察到,C类伤害性神经元Ih的增加主要与超极化激活的环核苷酸门控阳离子通道亚型2功能相关。这项研究是对C类伤害感受器参与穴位敏化神经机制的最直接证据。
根据响应特性的不同,C类伤害感受器可以进一步分为机械响应、热响应、冷响应等亚型。其中,C类伤害感受器中有一类具有“沉默”特性的亚型 ,在人体皮肤中的分布比例约为15%~20% ,与穴位的动态激活特性十分相似。生理情况下,这种沉默型C类伤害感受器对机械刺激不响应,而当组织损伤所致的病理条件下,这类感受器能够被“唤醒”,转换为机械响应性。如Feng等通过神经单纤维记录法,在小鼠结肠炎模型中观察到,造模后支配远端结肠的机械非敏感型感受器的比例从27%降至13%,而对机械刺激响应的C类感受器数量从34%增加到53%,提示至少有14%的沉默型伤害感受器在造模后被“唤醒”,转变为机械敏感型。这种转变的意义在于增加/放大伤害性信息向中枢的传递,提高机体对已有或潜在损伤的响应性,对于保护本体或促机体恢复具有重要意义。
反观穴位的起源,与疾病时古人对体表特定部位的按压、抓、摩等本能行为相关。生活中各种因素引起体表或内脏出现痛觉异常时,我们本能地会寻求或使用摩擦、抓捏、按压等刺激方法改善症状。同时,在穴位定位时,古人强调“欲得而验之,按其处,应在中而痛解,乃其腧也”(《灵枢·背腧》),提示疾病时用于治疗的腧穴会出现痛觉异常及区域大小的变化情况。这些机械类刺激产生的治疗效果是否与“唤醒”的C类沉默型伤害感受器相关?本课题组目前以结肠炎大鼠为研究对象,通过免疫荧光实验的方法,在结肠炎引起的敏化穴位皮肤及相应节段的DRG中,均能观察到用烟碱型乙酰胆碱受体亚型3(CHRNA3)标记的C类沉默型伤害感受器的分布(图1),为C类沉默型伤害感受器与穴位敏化的神经机制之间的联系提供了可能性依据,下一步将开展更系统的研究,以探讨穴位的动态敏化特性是否与沉默型C类伤害感受器激活相关。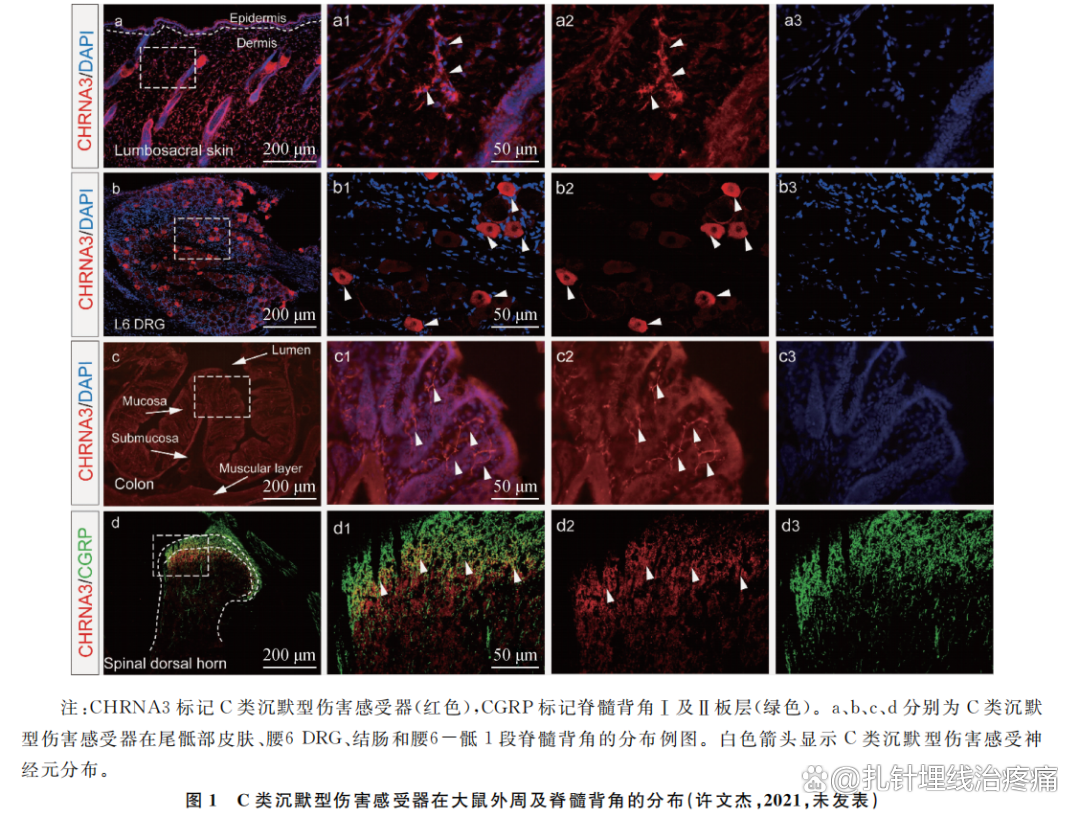
4C类伤害感受器介导穴位敏化时的病理变化——神经源性炎性反应
前期研究表明,神经源性炎性反应是敏化穴位的主要病理特征,与C类伤害感受器的激活直接相关。神经源性炎性反应主要是指当体表损伤或内脏器官受到缺血、牵拉、扩张等刺激时,可以激活Aδ与C类伤害感受器,并在其轴突末梢释放内源性炎性物质,如神经营养因子、缓激肽、组胺、前列腺素等,以及血管扩张物质CGRP与SP,激活局部的肥大细胞及免疫细胞,引起神经源性炎性反应。这些炎性物质构成“炎性池”,通过激活周围感受器上的相应受体(如激活Aβ感受器引起触诱发痛,激活C类沉默型伤害感受器增加机械响应强度),导致敏化范围扩大及痛觉敏化程度的加剧。其中,CGRP与SP能够增加血管通透性,导致血浆蛋白渗出及水肿。如动物实验中观察到的伊文氏蓝渗出现象就与敏化穴位穴区的血管通透性增加相关。
在敏化穴位的生物学效应中,神经源性炎性反应无疑是增加针刺等体表刺激信息传入、提高机体响应水平的重要神经基础。Wu等观察到针灸刺激正常穴位可以出现与敏化穴位相同的组织细胞化学改变(神经源性炎性反应),即敏化穴位具有一定的“拟针灸”效应,而针刺干预可以进一步放大并强化敏化穴位与内脏的联系,以促进机体的自我修复机制。
C类伤害感受器介导穴位可塑性的中枢及外周机制
如前所述,神经源性炎性反应是穴位敏化的主要病理特征,因此,探讨C类感受器介导神经源性炎性反应的中枢及外周神经机制是揭示穴位可塑性的关键点。在外周水平,伤害感受器的功能性变化是影响穴位功能的第一步。无论是外源性的体表刺激或内源性的因素作用于伤害感受器,均会兴奋伤害感受器产生动作电位并转变为敏化态(激活),表现为激活阈值降低、对刺激的反应程度增加等。伤害感受器的这种变化能够增加自身向中枢传递信息的效能,为强化针刺敏化穴位产生的信息传递奠定关键基础。目前研究认为,神经源性炎性反应的外周机制与C类伤害感受器主导的轴突反射及背根反射(DRR)密切相关。这两种反射机制为伤害感受器形成“自我意识”提供了条件,使其能够决定是否进一步将信息上传至脊髓或脊髓上水平。伤害感受器的这种功能正是穴位功能可塑性的重要基础。因为这种特殊的“自我意识”调节能够引起机体对敏化穴位的关注,驱动个体对体表出现痛觉异常改变预警部位的重视,并本能性地对敏化部位进行按揉或寻求规范性治疗。不过也有学者进一步指出,机械痛觉的敏化与中枢敏化更相关,而外周敏化主要介导病理条件下的热敏发生。
1 轴突反射
轴突反射是指体表或内脏的伤害性刺激导致DRG中的神经元(伤害感受器的胞体)兴奋时,产生的动作电位不仅可以顺向传导至脊髓背角,同时也可以通过外周分叉的侧支途径逆向传导至其外周端,并在轴突末梢释放血管活性物质如CGRP和SP,以及其他的活性物质(如炎性因子),引起神经源性炎性反应(图2a),同时炎性物质能够激活C类沉默型伤害感受器,使其获得机械敏感响应性及更强的热响应性,参与痛觉敏化的过程。研究者们在动物及人体试验中证实,轴突反射主要与C类伤害感受器相关。Lewin等以大鼠为实验对象,观察到皮肤中的C类伤害感受器参与轴突反射引起的血管扩张性渗出;Fang等在结肠炎大鼠模型中观察到,腰6-骶1 DRG中的C类伤害性神经元存在同时支配内脏与体表的分叉现象,参与结肠炎引起的体表敏化。而Schmelz等在记录人体试验中观察到,C类沉默型伤害感受器与轴突反射引起的皮肤红晕现象直接相关。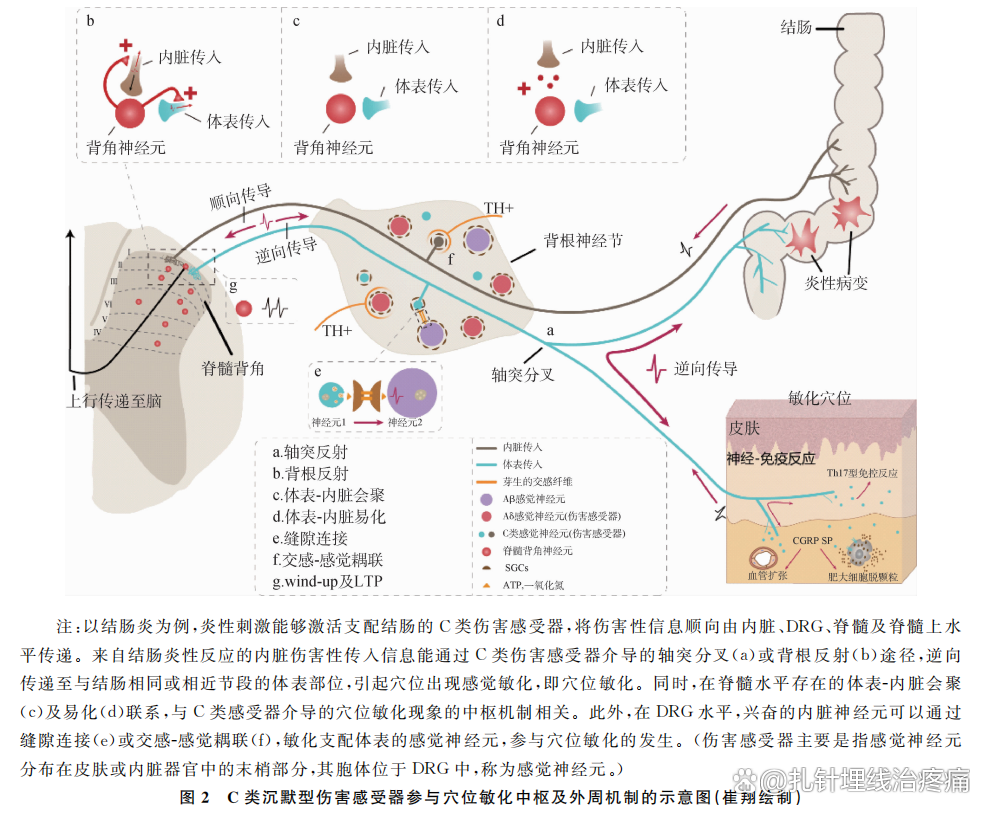
2 DRR
外周的伤害性信息激活伤害感受器并由其传入至脊髓背角,但脊髓背角也可以通过逆向的DRR动作电位将信息传递至外周端,释放SP、CGRP及其他炎性物质,导致并增强局部的神经源性炎性反应(图2b)。研究认为,足够强的初级传入纤维去极化(PAD)是引起DRR的必要条件,同时,PAD参与机体对运动行为控制的突触前抑制效应。PAD的产生及调制主要受初级传入纤维(尤其是C类纤维)及脊髓上水平(前庭核、脑干网状结构、红核、中脑导水管周围灰质、感觉运动皮层)的影响。已有资料显示,虽然大、中直径的Aβ、Aδ感受器及小直径的C类伤害感受器均与DRR相关,但DRR介导的神经源性炎性反应及触诱发痛的发生主要与C类伤害感受器兴奋相关。
3 会聚-易化学说
体表-内脏会聚及易化学说是牵涉性疼痛产生的基本机制之一,同时也是理解内脏疾病引起体表特定部位穴位出现敏化现象的理论依据。体表-内脏会聚主要是指体表和内脏伤害性信息传入在脊髓及脊髓以上水平发生会聚(图2c);而会聚易化学说则是指传递内脏感觉信息的传入纤维能够与接收体表信息的同一背角神经元形成突触联系,内脏疼痛性输入在脊髓背角形成一个“激惹灶”,易化正常来自躯体结构的信息,产生牵涉性疼痛表现(图2d)。目前认为,会聚易化学说与疾病时背角的中枢敏化有关。Luz等通过电生理实验观察到,来自体表与内脏C类纤维的轴突末梢可以与脊髓背角Ⅰ板层同一个投射神经元形成突触联系,证实了体表-内脏会聚/易化解剖学联系的存在。
4 DRG神经元的耦联激活机制
DRG是伤害性信息及本体感觉传入通路中的重要结构,是各种感受器的神经元胞体所在。目前研究认为,疾病时DRG的神经元能够出现耦联激活现象,其机制主要与卫星胶质细胞(SGCs)活化介导的缝隙连接(图2e),以及交感芽生引起的交感-感觉耦联相关(图2f)。在DRG神经元的周围分布着大量的SGCs,各种伤害性刺激时能够导致DRG神经元释放多种活性物质如一氧化氮、三磷酸腺苷(ATP)激活周围SGCs,通过缝隙连接影响临近的DRG神经元,使其出现自发性电活动;同时,DRG神经元释放神经生长因子等炎性物质,引起支配DRG神经元周围血管或分布于远端的交感节后纤维出现芽生现象,这些芽生的交感轴突末梢包绕DRG神经元,并释放去甲肾上腺素与ATP,敏化周围的神经元,最终导致DRG神经元的自发活动增强,增加伤害性信息向脊髓背角的传递,引起中枢敏化。
5 脊髓背角的中枢敏化
脊髓背角是整合外周感觉信息和执行脊髓上中枢调节指令的重要部位,脊髓水平的中枢敏化是继发性疼痛、触诱发痛和慢性痛的重要神经机制。大多数的Aδ与C类伤害感受器的中枢端投射到背角的Ⅰ~Ⅱ板层,其中,在背角的Ⅰ板层中存在着一定数量的投射神经元(PN),一部分PN能够将信息上传至丘脑、导水管周围灰质以及臂旁核中,另一部分将信息传递至脑干延髓头端腹内侧区,通过下行投射通路参与痛觉的产生及调制。目前认为,脊髓水平的中枢敏化机制与以下3点相关:①背角神经元的激活阈值降低、自发活动增强;②激活的C类伤害感受器介导的广动力神经元wind-up现象;③轴突的芽生。而背角神经元出现突触可塑性改变即长时程增强(LTP)是上述机制的重要基础。当外周的Aδ与C类伤害感受器被激活后,产生的动作电位能够顺行传递到轴突中枢端末梢,并释放CGRP、SP以及谷氨酸等物质。由于80%的投射神经元表达SP的受体神经激肽-1(NK1),因此,SP能够激活NK1+投射神经元,导致其出现wind-up现象;同时,谷氨酸能够与N-甲基-D-天冬氨酸受体结合,导致背角神经元与投射神经元出现wind-up现象及突触可塑性改变,如LTP(图2g),引起脊髓及脊髓上水平的中枢敏化,导致感受器的激活阈值降低,在相应节段部位出现痛觉过敏或触诱发痛现象。
近些年,有学者们在脊髓水平探讨了穴位敏化的中枢机制,并取得了一定的成果。
如Lv等以结肠炎大鼠为研究对象,通过痛行为检测观察到模型组大鼠出现足底机械痛及热痛敏现象;通过电生理记录刺激C纤维引起的背角场电位水平(反映背角神经元的LTP水平),结果显示模型组大鼠引起背角场电位的C纤维兴奋阈值较对照组大鼠显著降低,同时模型组背角的场电位显著增加,验证了内脏疾病引起的体表敏化与C类伤害感受器介导的背角神经元LTP变化相关。Rong等将芥子油注射到大鼠结肠造成结肠炎,通过电生理记录脊髓背角的广动力神经元活动,观察到芥子油注射后能够显著增加广动力神经元的放电水平,同时针灸体表部位能够兴奋广动力神经元的穴位感受野显著扩大,存在明显的穴位敏化状态(结肠炎引起的体表敏化程度)与针灸调节广动力神经元活动的量效关系。
总结与展望
穴位作为针灸学的关键角色,揭示其科学属性是推进针灸学科发展、促进针灸国际化水平的重要工作。近些年,与穴位相关的“泛穴现象”、穴位的特异性与非特异性作用、干针及其激痛点的定性等问题为针灸的研究工作带来重大挑战。其实,目前穴位研究中观察到的所谓“机械痛敏”“热敏”“压敏”仅是检测敏化穴位的一种物理方法,实质检测的都是感觉的敏感变化:Sensitization(敏化)!因此,穴位敏化的伤害感受器机制研究是揭示穴位敏化现象及其功能可塑性的重要切入点。虽然目前的研究已经观察到C类伤害感受器与穴位敏化的神经机制相关,但证据尚少,值得深入研究。作为C类伤害性感受的亚型,疾病时C类沉默型伤害感受器的“唤醒”特性使理解疾病时机体处理感觉信息的传入方式有了新的认识。需要思考的是,针灸、推拿等方法多属于机械类刺激,并且穴位敏化后有痛阈降低表现,是否疾病时被“唤醒”的C类沉默型伤害感受器与穴位的敏化特性及其功能可塑性直接相关?资料显示,C类沉默型伤害感受器具有更大的外周感受野,不仅分布于皮肤中,在内脏及关节中也有较高的分布比例(在心脏、膀胱、直结肠及膝关节的分布比例占到30%~90%不等)。近来,德国Vincenzo团队通过特异性标记C类沉默型伤害感受器,观察到沉默型伤害感受器主要为肽能的C类感受器,在颈1至骶1节段的DRG中均有大量分布,而它的中枢端主要投射至脊髓背角的Ⅰ板层,这里既是体表内脏神经元会聚易化的部位,同时也是投射神经元分布的主要位置,与穴位功能可塑性中枢神经机制密切相关。因此,结合C类沉默型伤害感受器生理及病理的不同响应特性,研究穴位敏化及其可塑性的神经机制,为揭开穴位原始属性的神秘面纱提供了新契机,也是揭示穴位敏化及其功能可塑性机制的新窗口。
